|
LA FATICA:
ASPETTI CENTRALI E PERIFERICI
Bisciotti GN. Ph D(1,2,3) ., Iodice PP (1),
Massarelli R (1). Sito
www. scienzaesport.com
1)
Dipartimento "Entraînement
et performance" , Facoltà di Scienze dello Sport, Università
Claude Bernard, Lione (F).
2)
Scuola Universitaria
Interfacoltà di Scienze Motorie, Torino (I)
3)
Consulente Scientifico
Internazionale FC
SdS.
Anno XXI, 54: 28-41, 2002
Key
words: Fatica periferica, fatica centrale, acidosi, fosfati inorganici,
eccitazione-contrazione, serotonina, ammoniaca.
Abstract:
Il fenomeno della fatica ha un’eziologia multifattoriale, non sempre
facilmente identificabile e la cui interpretazione spesso comporta
numerosi dubbi e non poche contraddizioni concettuali. Classicamente si
tende a suddividere il fenomeno in
fatica periferica e fatica centrale attribuendo alla prima cause
prevalentemente metaboliche ed alla seconda invece motivazioni
essenzialmente di tipo
neurale. Tuttavia il quadro generale non è sempre così perfettamente
distinguibile ed i vari fattori scatenati si sovrappongono molto spesso
in maniera indistinguibile, rendendo la situazione di difficile lettura
interpretativa. In questa breve review
si cerca di fare un punto sullo stato attuale di conoscenza della
problematica inerente l’insorgenza della fatica, sia periferica, che
centrale, sottolineando i molti punti di dubbio ed i possibili futuri
campi d’indagine.
INTRODUZIONE
Nel
corso degli ultimi trent’anni il concetto di fatica si è piuttosto
modificato ed in un certo
senso “evoluto”. Prima degli anni ’70 infatti, fisiologicamente
la nozione di fatica
era essenzialmente un sinonimo dell’esaurimento delle scorte
energetiche, prevalentemente dell’ATP e dell’ accumulo di sostanze
inibitrici nei confronti dei meccanismi di ripristino energetico (Westerblad
e coll., 1991). Solamente a partire dagli anni ’80 si è cominciato ad
interpretare il fenomeno
come multifattoriale e reversibile, considerando anche, sia la sua
diversa velocità, che i suoi differenti termini d’insorgenza. Più
tardi, a cominciare dagli anni ’90, si è potuto assistere ad un
crescente consolidamento dei concetti di plasticità muscolare, dei
meccanismi di ottimizzazione della produzione di forza da parte del
muscolo e della sua ricerca di “attivazione economica”, nonché più
recentemente all’apparizione del concetto di formazione di complessi
sub-cellulari tra i sistemi biologici e gli elementi ultrastrutturali
legato al concetto di formazione di micro-ambienti (Korge e Campbell,
1995).
Autori
come Korge e Campbell,
mettono in discussione il fatto che uno dei fattori scatenanti il
fenomeno della fatica possa essere costituito dalla mancanza di ATP, dal
momento che la deplezione di ATP viene efficacemente controbilanciata
dalla sua rigenerazione attraverso un fenomeno di down-regulation anche
nel muscolo affaticato. Potenzialmente questo fenomeno di
down-regulation potrebbe essere svolto da alcuni prodotti della reazione
ATPasica
come dall’accumulo di Pi od H+. I due Autori sottolineano
come esistano evidenze sperimentali che dimostrino che il legame della
creatin-kinasi e degli enzimi glicolitici nella vicinanza dei siti
d’idrolisi dell’ATP ed il loro accoppiamento funzionale con i
meccanismi di rigenerazione dell’ATP, potrebbe creare un
“micro-ambiente” che abbia un importante ruolo nella regolazione
della funzione ATPasica. Un’importante funzione in questo fenomeno di
rigenerazione dell’ATP può essere assunto dal valore ottimale di
ratio locale ADP/ATP, che sembrerebbe particolarmente importante nel
caso di un alto turnover dell’ATPasi. Sfortunatamente nel muscolo in vivo
non è dato conoscere il massimo rateo locale di rigenerazione
dell’ATP, in funzione della sua idrolisi, questa mancanza di
conoscenza di dati precisi è principalmente dovuta al fatto che, nella
determinazione in vitro questo valore viene di fatto
sistematicamente sottostimato.
Ed è proprio negli anni ’90 (Atlan
e coll., 1991) che appare per la prima volta il temine
anglosassone wisdom (che tradotto letteralmente significa
saggezza) che descrive il sistema di protezione di tipo progressivo
messo in atto dal muscolo contro il fenomeno della necrosi.
I
due principali processi implicati nell’insorgenza del fenomeno della
fatica, che sono costituiti dalla trasmissione del segnale nervoso e
dalla catena energetica metabolica, sono fortemente interagenti e si
sovrappongono costantemente, costituendo in tal modo, sia singolarmente,
che sinergeticamente, la causa scatenante della fenomenologia. La
bibliografia inerente la problematica fisiologica della fatica è
vastissima (si possono trovare oltre 3.800 articoli scientifici
sull’argomento) e costellata di numerose divergenze interpretative
riconducibili essenzialmente a problemi di standardizzazione e
riproducibilità tra i vari protocolli d’indagine. Soprattutto la
trasposizione di dati ottenuti in vitro, rispetto alla situazione
in vivo, è alquanto deludente. Occorre anche notare che, sia i
criteri di ordine biomeccanico, che quelli di tipo prettamente
biologico, inerenti il fenomeno della fatica, sono sovente mal definiti,
dal momento che occorre ricordare che la fatica, e conseguentemente la
sua modalità d’insorgenza, è “compito-specifica”, ossia presenta
una forte specificità nei confronti dell’attività che la ha indotta
(Allen e coll., 1995; Fitts e Metzger, 1993; Fitts, 1996; Green, 1997;
Mc Lester, 1997; Sejerstede
e coll., 1998; Westerblad e coll., 1991).
D’altro canto anche la classica forma iperbolica che descrive
il rapporto tra tempo limite e la percentuale
di forza massimale utilizzata nel corso dell’esercizio, ci sottolinea
l’aspetto fortemente multifattoriale del fenomeno (Rohmer, 1968).
In questo
lavoro prenderemo in esame i principali fattori che determinano
l’insorgenza della fatica periferica e
della fatica centrale per poterci meglio rendere conto della
complessità del problema e del suo aspetto “multiparametrico”, che
rende impossibile poter imputare ad un solo fattore l’insorgenza del
fenomeno.
LA
FATICA PERIFERICA
Come
già accennato il fenomeno della fatica è stato indagato tramite
l’utilizzo di svariati protocolli di studio, il più delle volte
difficilmente riproducibili e standardizzabili ed in ultima
analisi scarsamente confrontabili. Le metodiche
maggiormente utilizzate sono costituite da sperimentazioni su
muscolo isolato oppure in vivo, sia sull’animale, che sull’uomo. Un
grosso apporto metodologico è stato costituito, verso la fine degli
anni ’70, dall’avvento
della Risonanza Magnetica Nucleare (RMN) grazie alla quale si è reso
possibile lo studio non invasivo ed in tempo reale dei meccanismi
energetici cellulari, come ad esempio la concentrazione muscolare di
protoni. Grazie all’avvento di questa nuova tecnica si è potuto dare
vita a tutta una serie di modelli che hanno tentato e tentano di
descrivere, con la maggior precisione possibile, tutte le tappe che
portano alla produzione di forza all’interno del muscolo in attività.
Tuttavia, indipendentemente dalle tecniche di indagine utilizzate, il
concetto di fatica varia in funzione dei diversi Autori. Alcuni infatti
adottano come criterio valutativo delle variabili di tipo biomeccanico,
come la tensione muscolare, altri invece preferiscono adottare dei
parametri di ordine biologico come la concentrazione di alcuni composti,
oppure l’attività di alcuni enzimi o di alcuni complessi molecolari.
Da un punto di vista prettamente metodologico, i tre tipi di metodo di
lavoro maggiormente utilizzati nei protocolli d’indagine della fatica
muscolare sono costituiti dalla contrazione isometrica di tipo continuo,
dalla contrazione isometrica di tipo discontinuo e dalla contrazione
isotonica discontinua. Questa ultima modalità di lavoro può essere
svolta eccentricamente, concentricamente, attraverso la modalità
isocinetica oppure grazie ad una combinazione di queste differenti
possibilità. La durata, la
progressività e l’intensità delle esercitazioni proposte nei vari
tipi di protocollo utilizzati sono le più svariate e costituiscono un
ulteriore problema interpretativo.
I
MECCANISMI ED I SITI IMPLICATI NELL’ISORGENZA DELLA FATICA IL RUOLO
DELLE POMPE Na+/K+ ATPase E Ca++
ATPase
Come
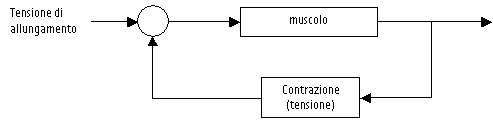
è noto il segnale chimico prodotto grazie all’acetilcolina, si
traduce a livello del sarcolemma nuovamente in segnale elettrico. Se
infatti una quantità sufficientemente elevata di questo
neurotrasmettitore si lega ad i recettori post-sinaptici, aumenta la
permeabilità del sarcolemma stesso nei confronti del sodio, da qui
risulta una depolarizzazione della membrana e la propagazione di un
potenziale di azione che si propaga lungo il sarcolemma. Questo
potenziale viene in seguito trasmesso ai tubuli traversi (sistema T)
verso l’interno della cellula. In questa sequenza di eventi sono
implicate, sia la pompa Na+/K+ ATPase, al livello
del sarcolemma, che la pompa Ca++ ATPase, a livello del
reticolo sarcoplasmatico. Le due pompe regolano i gradienti ionici
trans-membranari che sono necessari al fenomeno eccitatorio ed
all’attivazione dell’accoppiamento acto-miosinico. La pompa Ca++
ATPase presenta una forte specificità in rapporto ad i vari tipi di
fibra e ne condiziona la velocità di contrazione, al contrario la pompa
Na+/K+ ATPase presenta poche differenze in
rapporto alla tipologia delle fibre muscolari. Numerose sperimentazione,
effettuate su muscolo in vivo, dimostrano come il bloccaggio di queste
due pompe, causi un abbassamento della capacità di contrazione (Nielsen
e Harrison, 1998). Alla fine di un esercizio condotto ad esaurimento il
ritorno ad uno stato di omeostasi della pompa Na+/K+
ATPase, si presenta più rapido di quello relativo alla pompa Ca++
ATPase (Green, 1998), tuttavia l’alterazione della funzionalità della
pompa Na+/K+ ATPase, che si verifica in condizioni
di fatica, altera significativamente il gradiente membranario del
potassio (McLester, 1997). Durante gli esercizi prolungati, soprattutto
svolti ad una certa intensità, si verifica un’importante fuoruscita
di potassio, tale da essere notabile anche a livello della differenza
artero-venosa del catione stesso. La riduzione del potenziale di azione
che ne consegue, che è dell’ordine di circa il 50%, potrebbe essere
sufficiente per modificare la funzionalità dei tubuli traversi ed
impedire la liberazione di Ca++ dal reticolo sarcoplasmatico,
situazione che porterebbe ad una diminuzione della capacità di
produzione della forza da parte del muscolo (Nielsen e Overgaard, 1996;
Rios e coll., 1991; Rios e Pizzarro, 1988). La capacità di resistenza
contrattile, sembrerebbe quindi dipendere dall’efficienza della pompa
Na+/K+, anche se per alcuni Autori (Sjogaard,
1996), il ruolo del potassio nell’insorgenza della fatica dipenderebbe
dalla natura della sperimentazione (in vivo oppure in vitro)
e dall’intensità del lavoro imposto. In vivo ed a bassa intensità di
lavoro infatti il ruolo del potassio, nel fenomeno d’insorgenza della,
fatica sarebbe alquanto limitato, e l’apparizione quest’ultima
dipenderebbe essenzialmente ad una disfunzionalità del sistema T (Sjogaard,
1996). Al contrario nel caso in cui siano presenti un’alta
frequenza ed un importante intensità di contrazione, si verifica una
significativa elevazione del gradiente extracelluare di potassio che si
accompagna, sia ad una diminuzione del potenziale di membrana, che del
potenziale di azione e di velocità di propagazione dell’onda
elettrica (Sjogaard, 1996).
Questo
aumento del gradiente extracellulare di potassio, influenzerebbe
a sua volta il
fenomeno di retrocontrollo del debito sanguigno muscolare locale, la cui
conseguenza potrebbe essere la stimolazione di chemiorecettori arteriosi
che indurrebbero un aumento della pressione arteriosa (Paterson, 1996).
Il ruolo del potassio nell’insorgenza della fatica verrebbe
ulteriormente confermato dal fatto in numerose sperimentazioni, nelle
quali veniva aggiunto potassio nell’ambiente cellulare, si verificava
inequivocabilmente una diminuzione della produzione di forza, anche in
un muscolo inizialmente non affaticato (Sjogaard, 1996). Nelle
sperimentazioni in vitro, al contrario, non è possibile attribuire
al potassio un ruolo particolarmente importante nell’insorgenza
della fatica, a causa della sua diluizione immediata nell’ambiente
cellulare. Nelle esperienze in vitro, è piuttosto il calcio che
si presenta sempre ben correlato alla produzione di forza.
Tuttavia
occorre riportare come in bibliografia sia possibile ritrovare
protocolli sperimentali nei quali l’affaticamento sia sopravvenuto
senza che peraltro si potessero registrare significativi incrementi del
potassio extra-cellulare (Sjogaard, 1996), in alcune di queste
situazioni infatti il potenziale membranario e l’ampiezza del
potenziale d’azione rimanevano sostanzialmente stabili, anche in
presenza del fenomeno di affaticamento, e l’aumento del gradiente
extra-cellulare di potassio poteva addirittura influenzare positivamente
la produzione di forza. Questi dati ci fanno quindi chiaramente intuire
come il potassio non sia l’unico elemento responsabile della fatica
muscolare e come comunque non agisca sempre in modo diretto.
Altri
fenomeni che potrebbero essere collegati all’insorgenza della fatica,
riscontrabili nelle sperimentazioni in vivo, possono essere
costituiti dall’apparizione di sostanze antagoniste dell’acetilcolina
a livello della giunzione neuro-muscolare oppure dall’accumulo di
protoni.
IL
CALCIO INTRACELLULARE E L’ACCOPPIAMENTO ECCITAZIONE-CONTRAZIONE
All’interno
della cellula l’insorgenza della fatica sembra legata ad una
fenomenologia piuttosto complessa concernente sostanzialmente delle
variazioni, sia nella
distribuzione e nei legami, oltre che nella concentrazione e nei
movimenti del calcio (Williams e coll., 1995). Alcuni lavori (Westerblad
e coll., 1990) in effetti mostrano come dopo un periodo di contrazione
tetanica ad alta frequenza compreso
tra i 5 ed i 10 secondi, la concentrazione di Ca++ risultasse
minore al centro della fibra in rapporto a quanto non fosse al bordo
della fibra stessa , questo dato indicherebbe un deficit di liberazione,
al centro della fibra, probabilmente dovuto ad un difetto del potenziale
di azione del sistema T. Occorre a questo proposito considerare che, dal
momento che il sistema T non è solamente devoluto a veicolare il
potenziale d’azione ma anche ad indurre una sorta di
retroregolazione nei confronti dell’accumulo degli ioni calcio,
un loro cospicuo aumento potrebbe elevare la soglia di propagazione del
potenziale d’azione del sistema T stesso (McLester, 1997). Nonostante
tutto, comunque il ruolo del sistema T e del reticolo sarcoplasmatico
nell’insorgenza della fatica periferica, non è ancora del tutto
chiaro, anche se alcuni Autori (Chin e Allen, 1998; Linde e coll, 1998.)
concordano nell’associare la fatica con almeno tre meccanismi legati
al calcio di cui i primi due sono costituiti da una diminuzione, sia
della sua liberazione, che del suo ripompaggio da parte del reticolo
sarcoplasmatico ed il terzo è rappresentato da un abbassamento della
sensibilità delle miofibrille sempre nei confronti del Ca++
stesso. Anche il pH locale, come vedremo in seguito, potrebbe giocare
un qualche ruolo attivo in questo tipo di meccanismo. In definitiva
dunque tutte queste modificazioni riguardanti la concentrazione del Ca++
intracellulare, costituiscono una forte
causa di perturbazione del meccanismo di accoppiamento
eccitazione-contrazione, anche se occorre sottolineare che la diversa
tipologia delle fibre è differentemente influenzata da questa
catena di eventi. Le fibre ossidative infatti vengono meno perturbate
dalle variazioni della concentrazione di Ca++ intracellulare,
durante il loro ciclo di accoppiamento eccitazione-contrazione,
rispetto alle fibre glicolitiche (Stephenson e coll., 1998), questo
diverso comportamento potrebbe spiegare, almeno in parte, la loro
maggior resistenza alla fatica.
IL
RUOLO DELL’ ACIDOSI
Il
metabolismo dell’ATP è strettamente legato a quello dei protoni ed
all’equilibrio acido-basico del sarcoplasma (Sahlin, 1994;
Linderman e Gosselink, 1994). In effetti la quasi totalità delle
reazioni ossidative concernenti L’ATP, comprese la sua idrolisi e la
sua reintegrazione, vedono una liberazione ed un’assunzione di protoni
da parte dell’ambiente. L’idrolisi di una molecola di ATP libera un
protone, la glicolisi anaerobica citoplasmatica forma 2 molecole di ATP
per ogni molecola di glucosio utilizzata con la conseguente liberazione
di due protoni, come d’altro canto la glicogenolisi nella quale
vengono prodotte 3 moli di ATP per ogni mole di glicogeno e comunque
vengono rilasciati nell’ambiente cellulare 2 protoni. Il meccanismo
anaerobico alattacido, al contrario ha un bilancio protonico nullo,
essendo la scissione della fosfocreatina un meccanismo blandamente
alcalinizzante (Wooledge, 1998). A riposo, o nel
corso dell’esercizio svolto a bassa intensità, il sistema è
leggermente sbilanciato verso un modico accumulo di protoni, dal momento
che in simili condizioni, la loro produzione risulta maggiore, seppur
leggermente, del loro recupero, che si attua attraverso le vie di
resintesi dell’ATP. Il sarcoplasma riesce comunque a mantenere nel
corso del lavoro poco intenso, un pH relativamente stabile grazie, sia
all’intervento di numerosi sistemi tampone, che alla fuoruscita dalla
cellula di protoni e di gas carbonico. Questi sistemi sono
particolarmente efficaci, basti pensare che in totale assenza di tamponi
cellulari, il pH cellulare scenderebbe a 1.5 (Rouillon e Candau, 2000).
Alcuni di questi processi, come ad esempio il meccanismo di trasporto
dei bicarbonati, sono stati scoperti solamente di recente, tanto è vero
che nel 1994 Linderman e Gosselink sostenevano ancora l’impermeabilità
del sarcolemma nei confronti del bicarbonato. E’ facilmente
comprensibile dunque come, nonostante il loro indubbio interesse nei
confronti del fenomeno della fatica, questi aspetti siano ancora molto
inesplorati e poco conosciuti. Possiamo comunque dire che il potere
tampone del muscolo scheletrico risulta maggiore di quanto non sia
quello plasmatico ma minore di quello eritrocitario e che i principali
sistemi tampone sono costituiti dal sistema bicarbonato/acido carbonico,
dal sistema proteina/proteinato
e dal sistema fosfato monoprotonico/fosfato diprotonico. Dal momento che
negli esercizi di alta intensità la cui durata vada oltre qualche
secondo, la risentesi dell’ATP avviene essenzialmente tramite il
meccanismo anaerobico lattacido, la concentrazione di protoni
nell’ambiente cresce rapidamente, superando ben presto le possibilità
di controllo del sistema tampone , il risultato è un rapido
abbassamento del pH sarcoplasmatico (Mannion e coll., 1995). La
perfusione degli ioni H+ dalla fibra muscolare al torrente
circolatorio, avviene con una velocità di circa 30 volte maggiore
rispetto a quanto non sia per lo ione lattato (La-), questo
grazie alla loro minor dimensione (Shepard, 1986). Circa un terzo degli
H+ non sarebbe comunque associato agli La-, questo
starebbe ad indicare l’importante ruolo giocato in questo senso dal
sistema di scambio sodio/protoni e dai sistemi bicarbonato-dipendenti (Bangsbo
e coll., 1990). Il ruolo dell’abbassamento del pH nell’insorgenza
del fenomeno della fatica è un argomento molto indagato e per alcuni
aspetti controverso (Allen e coll., 1995;
Chin e Allen, 1998; Fitts
e Metzger, 1993; McLester, 1997; Westerblad
e coll., 1991) e l’elenco delle conseguenze fisiologiche che i vari
Autori attribuiscono all’acidosi è molto lungo: diminuzione
dell’attività della pompa sodio/potassio con conseguente apertura dei
canali potassici, diminuzione della fissazione del calcio sulla
troponina, dato il suo antagonismo con gli H+, diminuzione
nella formazione del numero di ponti acto-miosinici, diminuzione della
velocità di accorciamento, diminuzione dell’energia cellulare dovuta
ad un abbassamento dell’attività enzimatica principalmente della
fosfofruttochinasi, diminuzione della miosina ATPasi (che vede il suo pH
ideale situato a 7.2), diminuzione sia dell’uscita di calcio, che di
protoni dalla cellula, aumento della rigidità delle proteine. Tutto
questo quadro viene ulteriormente aggravato in condizioni
d’ipertermia. Numerosi studi confermano il ruolo effettivo sostenuto
dall’acidosi muscolare nell’insorgenza della fatica nel corso di
esercizi svolti ad alta intensità e di media durata (Linderman e
Gosselink, 1994). D’altro canto una controprova indiretta
dell’importanza dell’abbassamento del pH muscolare nel fenomeno
della fatica, è costituito dall’aumento della massa muscolare stessa,
e quindi del potere tampone del muscolo,
in seguito ad alcuni tipi di allenamento (Juel, 1998). Tuttavia
sono molte le perplessità espresse in merito all’abbassamento del pH
come maggiore responsabile della situazione di affaticamento periferico.
Quelle che potremmo definire come “divergenze interpretative”,
vanno dalla messa in discussione dei metodi d’indagine
utilizzati, come ad esempio il fatto che nella fibra isolata i protoni
escano più velocemente di quanto non facciano nella fibra in vivo, sino
alla critica di alcuni aspetti maggiormente specifici. Secondo alcuni
Autori il lattato non sarebbe né il solo, né tanto meno il principale
fornitore di protoni nel corso dell’esercizio muscolare. Secondo i
dati riportati da Sahlin (1992) il pH riscontrato su di un prelievo
bioptico muscolare in
condizioni di riposo e di fatica, passa da un valore di 7.1 a quello di
6.6, in queste condizioni la concentrazione di lattato aumenta, passando
da 1 a 30 mmol . l-1 di acqua intracellulare.
Contemporaneamente la degradazione di PCr e di ATP prima in ADP e
susseguentemente in AMP
provoca la liberazione di grandi quantità di acido fosforico (H3PO4)
che vede aumentare la sua concentrazione da 17 a 49 mmol
l-1 di acqua intracellulare. Sapendo che il pH
rappresenta il logaritmo decimale su base negativa (ossia
dell’inverso) della concentrazione di protoni, possiamo calcolare, sia
la concentrazione di protoni prima dell’esercizio (79 nmol
l-1 : 1 nmol = 10-6 mmol), che quella
riscontrabile dopo l’esercizio stesso (251 nmol
l-1): In quest’aumento di concentrazione di protoni
post-esercizio, che è
quindi di 172 nmol l-1,
il contributo della degradazione dei composti fosforici ad alta energia
(PCr, ATD, ADP) sarebbe di 1.5
volte maggiore rispetto a quello del lattato
(Sahlin, 1992).
Sempre
Sahlin (1992) ed altri Autori (Hirvonen e coll., 1987; 1992),
farebbero osservare come durante un esercizio di breve durata
svolto ad alta intensità, la degradazione di PCr e l’accumulo di
lattato siano tra loro in un rapporto molto vicino, se non eguale, ad
1:1. In altre parole
questi Autori sottolineano il fatto che quando la concentrazione di
lattato aumenta di 1 mmol l-1,
quella di PCr diminuisce di altrettanto. Come è noto La formazione di
una mole di creatina, proveniente dalla degradazione di una mole di PCr,
permette l’eliminazione di una mole di protone
e di una mole di lattato, in tal modo la degradazione della PCr
tamponerebbe la gran parte dei protoni forniti dalla glicolisi
anaerobica. Se è indubbio
che la contrazione muscolare, effettuata al di là di una certa intensità,
provochi un abbassamento del pH, che è a sua volta
responsabile dell’inibizione della PFK (fosfosfofruttochinasi),
l’enzima chiave della glicolisi lattacida ,
è altrettanto vero che il fenomeno che interagisce tra questi
due fattori, acidificazione dell’ambiente ed inibizione della PFK, non
è sempre perfettamente chiaro. L’attività ottimale dell’enzima PFK
si trova ad un pH di 7.03, ossia molto vicino al pH che presenta il
muscolo a riposo, la caduta del pH sino a valori di 6.63, livelli
d’altro canto facilmente raggiungibili nel corso di un’esercitazione
intensa, vede in effetti abbassarsi praticamente a zero l’azione
dell’enzima chiave del meccanismo glicolitico. Tuttavia anche in
simili condizioni, un certo numero di composti, che sono presenti a
livello muscolare, sono in grado di
rimuovere l’azione inibitrice svolta dai protoni sulla PFK
stessa. Ad esempio ad un pH pari a 6.63, l’aggiunta di fosfato
inorganico, sino al raggiungimento del livello di 20 mmol . l-1,riporta
l’attività enzimatica della PFK a circa il 40% delle sue capacità
(assumendo come 100% dell’attività della PFK quella registrabile a pH
7.3). Anche l’aggiunta di ADP, in ragione di 0.5 mmol l l-
riporta l’attività enzimatica della fosfofruttochinasi al 55% delle
sue piene potenzialità, mentre queste ultime rimontano sino a ben il
70% dopo l’aggiunta di una pur scarsa quantità di fruttosio 1,6
bifosfato, composto quest’ ultimo, che sappiamo essere ben presente
nel muscolo in attività, nel quale svolge peraltro degli importanti
ruoli di regolazione (Perronet, 1994). Questi dati potrebbero quindi
inficiare la teoria secondo la quale l’accumulo di lattato, ed il
conseguente abbassamento del pH, inibirebbero l’attività della PFK, o
per meglio dire, sottolineerebbero il fatto che tale fenomeno sia
perfettamente osservabile in sperimentazioni effettuate su muscolo
isolato ma non altrettanto evidente sul muscolo in vivo, nel quale il
livello di ATP e di fruttosio 6 fosfato ( da cui per fosforilazione da
parte dell’ATP deriva il fruttosio 1,6 bifosfato) sono molto più
elevate.
Anche
il ruolo dell’abbassamento del pH sull’apparato contrattile potrebbe
essere comunque messo in discussione. E’ conoscenza comune
l’antagonismo esistente tra i protoni e gli ioni calcio sui siti di
legame della troponina. Questo sarebbe in effetti un meccanismo di
difesa del muscolo che inibendo appunto il meccanismo di contrazione in
presenza di un pH eccessivamente basso, previene i possibili danni che
potrebbero conseguire al perdurare della contrazione muscolare in un
ambiente estremamente acido. Tuttavia se si osserva l’andamento
durante la fase successiva ad un esercizio muscolare intenso del
ripristino del pH e delle capacità contrattili del muscolo, possiamo
quantomeno mettere in dubbio le affermazioni precedenti. Dopo una
contrazione di tipo isometrico, mantenuta sino a quando la forza
espressa non cada al 50% della forza massimale volontaria, si registra
in effetti un pH molto basso (6.1-6.6), durante la fase di recupero il
pH ritorna verso valori normali solamente in tempi molto lunghi
dell’ordine di circa 10 minuti, inoltre durante i primi 2’ circa
della fase di recupero il pH continua ad abbassarsi, nonostante
l’interruzione del lavoro fisico, questa ulteriore
acidificazione dell’ambiente muscolare è dovuta alla liberazione di
protoni che avviene durante la resintesi della creatina in PCr.
Nonostante il fatto che il ritorno del livello del pH verso i valori di
riposo sia un processo relativamente lento, dell’ordine come abbiamo
detto di una decina di minuti, il muscolo riesce a ristabilire le sue
capacità contrattili in un arco di tempo molto più breve. Infatti dopo
il mantenimento di una contrazione isometrica che prosegua sino a che la
forza generata non cada a valori pari al 50% della massima forza
isometrica, il muscolo recupera completamente, ritornando quindi in
grado di generare nuovamente la stessa forza, dopo un periodo di riposo
compreso tra i 2 ed i 3 minuti (Sahlin e Ren, 1989). Nella corso dello
stesso studio i due Autori notarono come dopo 2 minuti di recupero la
diminuzione del livello degli H+ dovuta alla metabolizzazione
del lattato, fosse completamente controbilanciato dal rilascio di altri
H+ dovuti alla resintesi della PCr. Il fatto quindi che il
muscolo possa ritrovare le sue piene capacità contrattili anche in
mancanza di un innalzamento del pH, getta numerosi dubbi sull’assunto
secondo il quale l’abbassamento del pH, dovuto all’accumulo di
lattato, sia il responsabile dell’inibizione contrattile del muscolo e
quindi il responsabile dell’insorgenza del fenomeno della fatica
periferica. Per cui, anche se in effetti la fatica muscolare appare in
presenza di un abbassamento del pH, le evidenze sperimentali, in tutto
rigore, escluderebbero un rapporto di linearità tra pH e forza e/o pH e
fatica e non permetterebbero di andare al di là di una relazione di
coincidenza tra i due fenomeni in causa (Chin e Allen, 1998). D’altro
canto in bibliografia esistono numerosi esempi di sperimentazioni che
riferiscono l’insorgenza del fenomeno della fatica anche senza il
verificarsi di acidosi muscolare (per una review vedi Allen e coll.,
1995) Alcuni Autori infine proporrebbero di considerare la fase del
recupero in due periodi distinti, il
primo dei quali considerabile come “periodo di recupero rapido”,
sarebbe caratterizzato da un veloce ritorno verso lo stato basale del
meccanismo di accoppiamento eccitazione/contrazione e di regolazione del
calcio e risulterebbe pH-indipendente, ed un secondo, più lento, che
sarebbe, almeno in parte, legato al ritorno verso i valori basali, sia
dei protoni, che dei fosfati (Fitts e Metzger, 1993). Tuttavia occorre
comunque ricordare che alcuni studi
recenti svolti su modello
animale, nei quali una perfusione di La (con concomitante mantenimento
del pH a valori basali) faceva registrare una diminuzione della forza
contrattile ( Hogan e coll, 1995), potrebbero riaprire il dibattito sul
ruolo del lattato nell’insorgenza della fatica. Per giustificare
questi risultati gli Autori avanzavano l’ipotesi dell’aumento della
forza ionica che sarebbe la responsabile di un’alterazione nella
formazione dei ponti acto-miosinici. In questo senso va anche un'altra
sperimentazione, sempre effettuata su modello animale, nella quale gli
Autori concludono che l’aumento di La possa avere un effetto negativo
sulla capacità di produzione di forza da parte del muscolo,
probabilmente a causa di un meccanismo d’inibizione nei confronti del
rilascio di Ca++ da parte del reticolo sarcoplasmatico (Stephenson
e coll., 1998).
Il
ruolo dei fosfati inorganici nella forma mono e diprotonica
I
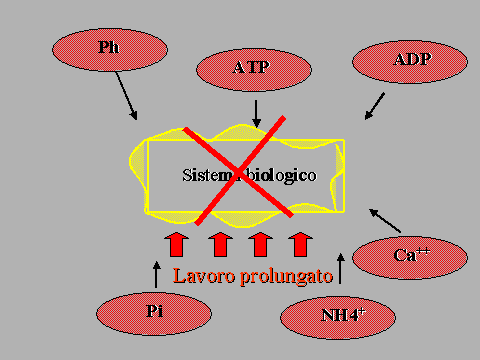
fosfati inorganici (Pi) sono dei
metaboliti derivanti dall’idrolisi dell’ATP e della PCr e la loro
concentrazione tende ad aumentare nell’ambiente indipendentemente
dalla durata dell’esercizio svolto. In bibliografia si possono
ritrovare numerosi lavori che tendono ad evidenziare il ruolo svolto da
queste sostanze nella diminuzione della prestazione fisica legata
all’insorgenza della fatica, oppure all’ischemia od all’ipossia.
L’aggiunta di poche millimoli di Pi nell’ambiente muscolare induce
una diminuzione della capacità contrattile, con una conseguente
diminuzione della produzione di forza ed un
cambiamento , sia del ciclo oscillante, che dell’attività
ATPasica. Vale la pena soffermarci a questo punto sul meccanismo del “
ciclo oscillante”: quando le fibre muscolari oscillano a 5-15 Hz per
circa il 2% della loro lunghezza di riposo, sono in grado di produrre
un’importante potenza meccanica idrolizzando praticamente il doppio
quantitativo di ATP, per unità media di tensione, rispetto alla condizione statica.
L’attività ATPasica risulta correlata
linearmente alla tensione
media prodotta durante l’oscillazione. L’aggiunta di Pi o di solfato
riduce, sia il costo della tensione, che la frequenza ottimale di
oscillazione di lavoro perturbando in tal modo il sistema (Pybus
e Tregear 1975). I Pi sono presenti a livello organico in due forme: la
prima monoprotonica e la seconda diprotonica la cui proporzione di
presenza dipende dalla
concentrazione dei protoni presente nell’ambiente. Nel momento in cui
il pH cellulare si abbassa, pressoché la totalità del Pi presente
passa alla forma diprotonica. Questo passaggio del Pi dalla forma
monoprotonica a quella diprotonica è correlato alla diminuzione della
forza contrattile, mentre questa correlazione non si registra con la
forma monoprotonica. Questi effetti variano in rapporto alla tipologia
delle fibre con ogni probabilità in ragione della diversa sensibilità
che le fibre medesime presentano per ciò che riguarda la loro attività
ATPasica nei confronti del Pi. Nel caso di esercizi di breve durata
svolti ad alta intensità, il verificarsi di un rapido ed importante accumulo di Pi, dovuto al massiccio
intervento del meccanismo anaerobico alattacido, costituisce uno dei più
importanti fattori responsabili dell’insorgenza della fatica
muscolare, assumendo in questo caso una valenza ancor maggiore di quella
rivestita dai meccanismi di perturbazione ionica. Questo sarebbe
tuttavia in contraddizione con quanto riportato da alcuni studi (Greenhaff,
1995; Mujika e Padilla, 1997) che riferiscono come una supplementazione
di creatina possa, aumentando le scorte di fosfocreatina, ritardare
l’apparizione del fenomeno della fatica.
In effetti una supplementazione di creatina, generando attraverso
il fenomeno dell’idrolisi Pi, dovrebbe al contrario essere un fattore
inducente la fatica (Sahlin e coll., 1998.). Anche durante le
esercitazioni di lunga durata svolte a bassa intensità, il sistema dei
fosfageni, che in questo caso si trova accoppiato ad un forte fenomeno
di idrolisi dell’ATP, può comunque indurre un fenomeno di elevazione
della concentrazione di Pi tale da comportare il passaggio dei ponti
actomiosinici dal loro livello di alta produzione di forza a quello
basso, fenomeno che potrebbe essere alla base della teoria del sistema
oscillante proposto da Mc Lester (1997). L’allenamento potrebbe
giocare un ruolo sostanziale nell’incidenza dei fenomeni sopra
descritti, inducendo una progressiva tolleranza a concentrazioni di Pi
sempre maggiormente elevate (Mc Lester, 1997).
Il
ruolo dell’adenosindifosfato
Recentemente
alcuni ricercatori hanno rivolto la loro attenzione al possibile ruolo
svolto dall’adenosindifosfato (ADP) nell’instaurarsi del fenomeno
della fatica periferica, prendendo in considerazione anche il rapporto
ADP/ATP (Allen e coll., 1995; Mc Lester, 1997; Sahlin, 1998). Questo
composto, come sottolineato nel modello energetico proposto da Mc Lester
(1997), svolgerebbe un ruolo di primo piano nel meccanismo di
transizione dallo stato di bassa a quello di alta energia ed è inoltre
considerabile a tutti gli effetti come il principale responsabile del
distacco dei ponti acto-miosinici. Nel momento in cui la sua
concentrazione subisce un sostanziale aumento, l’ADP ostacola l’ ATP
nel meccanismo di distacco dei ponti actomiosinici, diminuendo in tal
modo la forza prodotta dal sistema oscillante. Questo effetto
d’inibizione sul distacco dei ponti verrebbe ulteriormente enfatizzato
in presenza di basse concentrazioni di ATP.
Il
ruolo dellA componente lenta del VO2 nel caso particolare
della corsa.
Durante
uno sforzo ciclico effettuato ad un
carico costante d’intensità inferiore a quella della prima
soglia ventilatoria,
soprattutto se svolto in posizione eretta come nel caso della corsa,
è possibile notare un primo repentino incremento del VO2,
essenzialmente ascrivibile al veloce aumento del flusso ematico
polmonare, che caratterizza la prima fase denominata “cardioritmica”,
alla quale fa seguito una seconda fase, caratterizzata da un aumento
meno ripido del VO2, legata all’arrivo del flusso ematico
proveniente dai distretti muscolari
attivi. Questa
seconda fase porta a sua
volta, in circa 3’, al
raggiungimento della terza fase, detta di stady-state, in cui il consumo
di O2 si stabilizza.
Nello
svolgimento di un lavoro effettuato ad un’intensità maggiore rispetto
alla prima soglia ventilatoria, la cinetica del VO2 cambia
sostanzialmente. In questo caso infatti alla seconda od alla terza fase,
si sovrappone una nuova componente caratterizzata da una cinetica più
lenta che, appunto per questa sua caratteristica, prende il nome di
“componente lenta del VO2”. La componente lenta del VO2
(cl VO2) rappresenterebbe quindi un “eccedenza” di VO2
che, sino a determinati carichi, consente il raggiungimento di uno
steady-state ritardato. Nel caso invece in cui
i carichi di lavoro siano particolarmente intensi, non diviene più
possibile il raggiungimento di uno stato di steady-state ed in tal caso
la cl VO2 concorrerebbe al raggiungimento del massimo valore
di VO2, valore peraltro superiore a quello prevedibile dalla
relazione VO2/carico sotto soglia (W) e quindi al rapido
raggiungimento dell’esaurimento da fatica. La cl VO2
comporterebbe un aumento del costo VO2/W, che passerebbe dai
circa 10 ml /W registrabili sotto-soglia, ai circa 12-13 ml/W
osservabili durante il lavoro sopra-soglia (Maione e coll., 20001),
evidenziando in tal modo una perdita di efficienza muscolare. La cl VO2
viene pressoché unanimemente spiegata dai diversi Autori, attraverso
fenomeni prevalentemente muscolari legati al progressivo reclutamento,
durante l’attività svolta ad alta intensità, di fibre di tipo II, il
cui rendimento è minore rispetto a quelle di tipo I. La cl VO2
quindi farebbe parte integrante del fenomeno della fatica e sarebbe une
delle principali cause, in attività come la corsa,
della progressiva riduzione dell’efficienza muscolare (Whipp e
Wassermann, 1970; Jacobsen e coll., 1998).
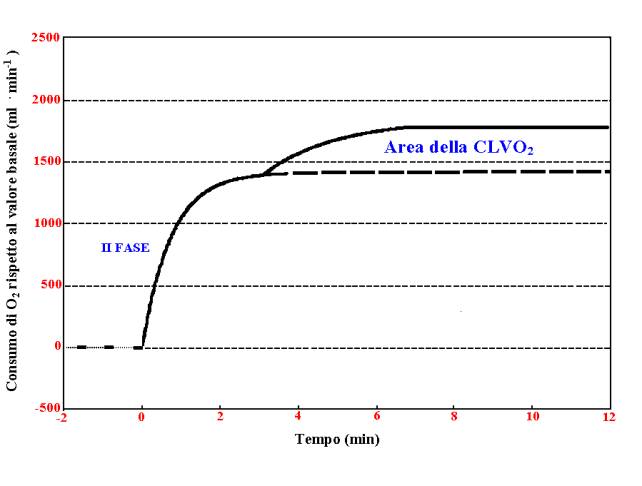
Figura
1 : la componente lenta del VO2.
LA FATICA CENTRALE
Con il termine di fatica nervosa o centrale
si intende tutto quel complesso di fattori che determinano la
diminuzione della contrattilità muscolare indipendentemente dai fattori
intramuscolari e/o metabolici. L’implicazione di fenomeni centrali
nell’insorgenza della fatica è dimostrata da alcune sperimentazioni (Bigland-Ritchie
e coll., 1979) che dimostrano come la stimolazione elettrica di un
complesso muscolare affaticato, permetta di recuperare una certa
percentuale del livello iniziale di forza. In questo tipo di
sperimentazioni alcuni soggetti furono sottoposti ad un lavoro di
tipo intermittente a carico del soleo, sino a che non fosse raggiunto un
livello di affaticamento tale da diminuire la forza del distretto
muscolare sino al 50% della massima capacità contrattile. Raggiunta una
simile situazione, gli Autori riferiscono di come l’imposizione di una
stimolazione elettrica permetta di ritrovare un livello di forza pari
all’80% del livello massimale, attribuendo in tal modo la differenza
tra i due valori alla fatica di tipo centrale. Occorre tuttavia notare
che il parziale recupero dei livelli iniziali di forza indotto dall’elettrostimolazione,
risulterebbe essere dipendente, sia dal gruppo muscolare considerato,
che dal tipo di esercitazione che ha causato la condizione
di affaticamento, senza dimenticare lo stato motivazionale del
soggetto (Guézennec, 2000). In ogni caso questi dati sottolineerebbero
come, in stato di affaticamento, il Sistema Nervoso Centrale (SNC) sia
incapace di generare uno stimolo adeguato (Bigland-Ritchie e coll.,
1979; Enoka e Stuard, 1992). Oltre a questo effetto sulla fatica acuta,
altri Autori hanno dimostrato come l’elettrostimolazione sia in grado
di permettere un parziale recupero dei livelli di forza persi in seguito
ad un fenomeno di fatica cronica come quello costituito dal
sovrallenamento (Bayley e coll., 1993). Tuttavia questo tipo di
sperimentazioni basate sulla contrazione elettroindotta, non riesce a
dimostrare appieno il ruolo che il comando nervoso, proveniente dal SNC,
ricopre nell’insorgenza della fatica, l’elettrostimolazione infatti
può indurre anche un potenziamento del comando nervoso periferico,
provocando in tal modo un aumento in toto del comando nervoso che
arriva a livello muscolare. Per questo motivo quindi la stimolazione
elettrica non può essere considerata come una tecnica specificatamente
rivolta all’indagine del fenomeno
della fatica indotta da un deficit di funzionamento del SNC. Non mancano
comunque esempi in letteratura che, proprio per dimostrare senza
possibilità possibili dubbi il ruolo del SNC nell’insorgenza del
fenomeno della fatica, hanno utilizzato come tecnica di studio la
stimolazione diretta della corteccia motoria. In quest’ambito uno
studio condotto da Maton sui primati (1991), utilizzando una tecnica di
registrazione dell’attività elettrica cerebrale dei neuroni della
corteccia motoria primaria, tramite un impianto elettrodico intracranico,
dimostrò come la contrazione esaustiva del bicipite brachiale
comportasse una diminuzione dell’attività elettrica dei neuroni
considerati. Il ruolo ricoperto dalla corteccia motoria primaria
nell’insorgenza della fatica, è stato confermato in seguito anche
nell’uomo grazie all’utilizzo di una tecnica non invasiva,
costituita dalla stimolazione della corteccia primaria tramite dei campi
magnetici intensi (Gandevia e coll., 1996). Attraverso questo studio gli
Autori hanno potuto dimostrare come la superimposizione di una corrente
magnetica transcranica permetta di diminuire parzialmente gli effetti
che la fatica provoca sulle possibilità di mantenimento della forza
contrattile. Tuttavia occorre notare che una parte degli effetti della
fatica non può comunque essere spiegata attraverso l’utilizzo di
queste tecniche. In ogni caso tutte le sperimentazioni che si basano
sulla stimolazione, effettuata a diversi livelli del tratto nervoso,
permettono di formulare la verosimile ipotesi dell’esistenza
di una fatica di ordine centrale, evidenziabile attraverso una
diminuzione del comando nervoso preposto alla contrazione muscolare,
anche se il ruolo dei fattori di ordine metabolico periferico giocano un
ruolo predominante per ciò che riguarda la diminuzione delle capacità
contrattili muscolari. Inoltre alcuni aspetti della fatica di ordine
centrale restano ancora da chiarire completamente, come ad esempio il
fatto che il fenomeno sia riconducibile ad un meccanismo inibitorio che
si presenterebbe a livello di alcuni gruppi di neuroni, oppure sia
piuttosto costituito da un fenomeno inibitorio più generalizzato,
causato dai dei meccanismi che agiscono a livello globale sull’insieme
delle funzioni nervose. La risposta a questo tipo di domanda non è
ancora del tutto chiara, anche se le attuali teorie neurochimiche della
fatica sembrerebbero propendere maggiormente per la seconda ipotesi (Guezennec,
2000).
GLI ASPETTI NEUROCHIMICI DELLA FATICA CENTRALE
L’evidenza del ruolo della fatica centrale,
comprovato attraverso le varie sperimentazioni di tipo
elettrofisiologico, ha spinto numerosi Autori a formulare differenti
ipotesi sul ruolo svolto dai neuromediatori centrali nel corso
dell’esercizio esaustivo (per una review vedi Meeusen e coll., 1995).
Tutti gli studi rivolti a questo particolare aspetto del fenomeno,
dimostrano ampiamente coma la fatica induca, sia nell’uomo, che
nell’animale, un cambiamento del modello comportamentale (Dishman,
1997). Nell’ animale si può sostanzialmente notare una diminuzione
delle attitudini comportamentali rivolte alla vita di relazione, mentre
nell’uomo le risposte sono maggiormente complesse e sembrano dipendere
dal tipo di attività responsabile del fenomeno di affaticamento.
Possiamo tuttavia in linea generale osservare nel modello umano, come
conseguenza ad un esercizio di tipo esaustivo, una diminuzione delle
capacità decisionali, sia per quello che riguarda la capacità di presa
d’informazione, sia per ciò che concerne l’interpretazione dei
segnali visivi (Koutedakis, 1995), nonché una diminuzione della memoria
a corto termine (Guézennec, 2000). Inoltre la fatica cronica può
essere all’origine di uno stato ansiogeno o depressivo (Dishman,
1997). Anche la secrezione di catecolamine potrebbe giocare un ruolo
importante negli aspetti di natura neurochimica legati alla fatica
centrale. Alla fine di un esercizio
esaustivo è infatti possibile notare nel tessuto cerebrale una
diminuzione della concentrazione di noradrenalina, che sembrerebbe
essere dovuta ad un aumento del suo turn over. Lo stesso fenomeno
è osservabile, in maniera ancor più evidente, a livello del tronco
cerebrale, dell’ippocampo e dell’ipotalamo (Gandevia e coll., 1996).
Questo quadro sarebbe simile a quello osservato nel corso di stress
psicologico acuto nel quale è appunto possibile notare una deplezione
delle riserve di catecolamine cerebrali. La conseguenza di questa
diminuzione nelle riserve di noradrenalina
si ripercuoterebbe a
livello comportamentale e potrebbe essere responsabile dell’insorgenza
di possibili stati depressivi.
Rimanendo nell’ambito della risposta
adrenergica, è importante notare che anche la dopamina può influenzare
fortemente l’attività muscolare. L’aumento dell’attività
dopaminergica nello striatum
induce infatti un aumento spontaneo della motricità.. A livello
cerebrale, durante un esercizio prolungato è possibile osservare in un
primo tempo un leggero aumento della concentrazione dopamina, a cui fa
seguito, nella seconda parte dell’esercizio stesso, quando
quest’ultimo si avvicina al punto di esaurimento, una sua
leggera diminuzione (Seguin e coll., 1998). Questa variazione
della concentrazione di dopamina cerebrale nel corso di un esercizio
prolungato che porti ad esaurimento, potrebbe essere giustificata dal
fatto che questo neuromediatore attraversi una prima fase accelerata di
liberazione a cui consegua una seconda fase di diminuzione secretoria
dovuta ad un esaurimento delle sue riserve neuronali. Sempre a questo
proposito è stata avanzata l’ipotesi di una deplezione di tirosina
che costituisce il precursore delle catecolamine. In qualsiasi caso il
ruolo ricoperto dalle catecolamine nell’insorgenza
dell’affaticamento organico, è sperimentalmente provato dal fatto che
la somministrazione, prima dell’esercizio, di anfetamina e di agonisti
dopaminergici e/o adrenergici, aumenta nell’animale la durata di
sopportazione allo sforzo (Seguin e coll., 1998).
IL RUOLO DELLA SEROTONINA NELL’INSORGENZA DELLA FATICA
Il ruolo svolto dalla serotonina nel fenomeno
della fatica organica, è ormai divenuto un aspetto dogmatico del
problema in seguito soprattutto ai lavori di sintesi svolti da
Newsholmes e coll. (1987) e Chauloff (1989). Quest’ultimo fu il primo
a mettere in evidenza sperimentalmente l’aumento della concentrazione
di serotonina a livello
cerebrale in seguito ad esercizio prolungato e/o ad allenamento inteso
protratto per più settimane. D’altro canto numerosi altri lavori,
anche se non in ambito prettamente sportivo, avevano già sottolineato
il ruolo della serotonina sul sonno, l’assunzione alimentare, gli
stati ansiosi e quelli depressivi.
A questo proposito ad esempio possiamo ricordare come sia da tempo noto
che uno stato ansioso sia caratterizzato da un aumento della
concentrazione cerebrale di serotonina, mentre al contrario, uno stato
depressivo veda ridursi i livelli di serotonina cerebrale. Per tutta
questa serie di motivi il fatto che all’esercizio prolungato che
conduce al fenomeno della fatica, consegua un aumento dei livelli di
serotonina cerebrale, ha portato alla
naturale formulazione dell’ipotesi che questo neuromediatore sia
fortemente implicato, se non addirittura il responsabile
principale, dell’insorgenza della fatica centrale. In questo senso
numerose sperimentazioni condotte su modello animale confermano il fatto
che l’aumento o la diminuzione del tono serotoninergico, indotto
attraverso l’utilizzazione di agonisti od antagonisti
serotoninergici, influiscano sul fenomeno d’insorgenza della
fatica (Bailey e coll. 1993). Occorre comunque ricordare che lo stesso
tipo di sperimentazioni condotte sull’uomo, non ha permesso di
confermare i risultati ottenuti sull’animale (Seguin e coll., 1998).
Altre sperimentazioni, sempre effettuate allo scopo di poter confermare
il ruolo svolto dalla serotonina sulla comparsa insorgenza della fatica,
hanno utilizzato la somministrazione di aminoacidi a catena ramificata (AABC).
Gli AABC infatti entrando
in competizione con il triptofano, sostanza precursore della serotonina
a livello del passaggio attraverso la barriera emato-encefalica,
dovrebbero limitare la produzione di quest’ultima a livello cerebrale.
Tuttavia i risultati ottenuti da Blomstrand e coll., (1991), dopo la
somministrazione di AABC prima di una prova di maratona, hanno permesso
di evidenziare solamente un incremento dei risultai ottenuti nel corso
di una batteria di test psico-sensoriali, ma non un incremento della
prestazione di gara. Questi risultati sono in linea con quelli ottenuti
da Bigland-Richie e coll.(1979), i quali non riportano di alcun
miglioramento della performance, durante un raid effettuato in alta
quota, in seguito alla somministrazione di AACB. L’insieme delle
sperimentazioni condotte sull’uomo quindi non permetterebbe di
evidenziare nessun risultato positivo, in termini di incremento della
performance, legato alla diminuzione del fenomeno di affaticamento,
indotto dall’utilizzo di AABC. Ma vi sono
numerosi altri limiti e contraddizioni nella teoria che vede la
serotonina come principale fattore d’insorgenza della fatica centrale,
uno di questi è costituito dalla mancanza di coerenza tra i dati
desumibili dai test comportamentali e gli effetti psicotropi
attribuibili all’azione della serotonina stessa. La fatica acuta od il
sovrallenamento cronico, infatti , indurrebbero l’insorgenza di turbe
comportamentali a sfondo principalmente depressivo caratterizzate da una
carenza serotoninergica (Guèzennec, 2000). Questi dati mal si
concilierebbero con l’iperserotoninergia che si registra nel corso
dell’esercizio fisico strenuo. Seguin e coll. (1998) hanno tentato di
spiegare questa contraddizione mettendo in evidenza una diminuzione
della recettività di alcuni recettori serotoninergici in seguito
all’esercizio prolungato. D’altro canto anche altri risultati molto
recenti riferirebbero di una caduta, al di sotto dei livelli basali,
della concentrazione di serotonina in alcune aree cerebrali,
riscontrabile alla fine dell’esercizio (Guèzennec, 2000).
I due dati di cui sopra, potrebbero quindi far propendere verso
un ipotesi di carenza di tono serotoninergico
che sopravverrebbe durante la fase di recupero successiva
all’esercizio. In tutti i casi la mancanza di omogeneità e di
perfetta coerenza tra i vari risultati ritrovabili in letteratura
mostrano come, in ultima analisi, sia sostanzialmente erroneo limitare
il fenomeno dell’insorgenza della fatica centrale esclusivamente alla
teoria serotoninergica. In effetti gli aspetti comportamentali, ivi
compreso quindi il fenomeno della fatica, risultano essere influenzati
per la maggior parte dei casi, da un delicato equilibrio esistente tra
numerosi neuromediatori. A titolo di esempio possiamo ricordare come gli
studi inerenti l’aspetto neurochimico del sonno, mostrino come
quest’ultimo dipenda da una complessa azione sinergica ed
interdipendente di numerosi neuromediatori in altrettanto numerosi
ambiti strutturali, gli aspetti neurochimici della fatica quindi
potrebbero essere improntati su di un meccanismo del tutto, od in parte
simile a questo.
IL RUOLO DELL’AMMONIACA
Occorre
anche ricordare il possibile meccanismo d’intervento dell’ammoniaca
legato alla manifestazione della fatica. L’encefalo utilizza come via
di metabolizzazione dell’ammoniaca la trasformazione del glutammato in
glutammina. L’iperammonemia quindi provoca una diminuzione della
concentrazione del glutammato in alcune aree cerebrali specifiche. Dal
momento che il glutammato costituisce il principale precursore
dell’acido gamma amminobutirrico (GABA), questa catena di eventi porta
ad un abbassamento della sua concentrazione a livello encefalico. Il
GABA è il neurotrasmettitore maggiormente presente a livello del SNC ed
esercita un importante ruolo di regolazione, di tipo inibitorio, sulla
liberazione di altri neurotrasmettitori, oltre ad agire direttamente sui
nuclei grigi della base facilitandone il compito di regolazione che
questi svolgono sulla motricità. La carenza di GABA inoltre gioca un
ruolo fondamentale nella patogenesi di alcune malattie come il morbo di
Parkinson e la Corea di Huntington. Tutta questa serie di dati farebbe
ragionevolmente propendere verso l’ipotesi che la carenza del sistema
GABAenergico, registrabile nel corso dell’esercizio intenso e
prolungato, possa giocare un ruolo importante nella manifestazione della
fatica a livello centrale. I risultati sperimentali sembrerebbero
confermare, per lo meno parzialmente, questa ipotesi, mostrando come il
sistema GABA-glutammato-glutammina
sia particolarmente attivo, in alcune aree cerebrali, nel corso
dell’esercizio esaustivo.
UN
MODELLO TRIDIMENSIONALE DELLA FATICA.
Secondo
alcuni autori il rapporto tra l’intensità dello sforzo e la
sensazione di fatica può essere anche interpretato attraverso tre
modelli fortemente interagenti tra loro.
Il
primo modello è il già descritto modello “classico” della fatica
periferica, denominato appunto “modello periferico”, nel quale i
fattori regolatori e/o inibitori sono esclusivamente di ordine
metabolico (Kay e coll., 2001; Kirkendall, 1990; Fitts, 1994; Basset
e Howley, 1997.).
Nel secondo modello, denominato “centrale-teleoanticipatorio” , il
meccanismo di controllo della fatica funziona essenzialmente come un
dispositivo di sicurezza che viene posto in atto da un meccanismo
subcosciente a livello cerebrale. Il meccanismo regolatorio
cerebrale viene modulato sulla base di input, sia centrali, che
periferici, ed il cui scopo è quello di preservare l’integrità
strutturale della fibra muscolare, prevenendo possibili danni
irreversibili a quest’ultima attraverso una riduzione od un arresto
totale dell’attività. Nel modello “centrale-teleoanticipatorio”,
il cervello svolge il ruolo di principale regolatore dell’intensità e
della durata dell’esercitazione, che viene mantenuta ad un grado
sub-massimale prefissato in modo
tale che il sistema periferico non sia mai utilizzato a livelli
massimali (St
Clair Gibson e coll., 2001; Wagenmakers, 1992; Kay e coll., 2001.)
In
questo secondo modello quindi il fenomeno della fatica può essere
considerato come un vero e proprio “atto anticipatorio di sicurezza”
che abbia lo scopo di prevenire, sia un eccessivo accumulo di metaboliti,
che un’esagerata deplezione di substrati energetici. In questo secondo
modello, l’attività non è mai massimale ma viene al
contrario mantenuta costantemente a livelli sub-massimali. Ulmer (1996)
avanza l’ipotesi che in questo modello i comandi neurali efferenti
regolino a livello del muscolo scheletrico, non solamente i pattern di
attivazione spaziali e temporali, ma anche il rateo metabolico
responsabile della produzione di potenza da parte del muscolo. Questo
tipo di meccanismo protettivo, potrebbe spiegare come nel muscolo
scheletrico la concentrazione di ATP non scenda mai al di sotto del
60-70% dei valori di riposo anche durante un esercizio di tipo esaustivo
(Fitts, 1994; Spriet e coll., 1987). Il modello “centrale-teleoanticipatorio”, sarebbe quindi il responsabile del
decremento dell’intensità dell’esercizio anche in presenza di
sufficienti riserve energetiche, per cui la manifestazione di fatica
sarebbe il risultato di un comando efferente di tipo inibitorio,
derivante da una sorta di “calcolo mentale”. In altre parole il
decremento dei comandi efferenti di origine neurale, sarebbe causato
dagli adattamenti a livello corticale del processo subcosciente teleoanticipatorio che si verificano in
seguito alle risposte agli input afferenti di origine metabolica
provenienti dagli organi e dalle strutture periferiche. Nel terzo tipo
di modello denominato di
“discussione-cognitiva”, è la sensazione di fatica stessa che, a
livello cosciente, utilizzando le antecedenti esperienze come secondo
termine di paragone, regola l’intensità dell’esercizio. In questo
terzo modello la fatica costituisce una sorta di processo continuativo
che modifica costantemente lo stato funzionale dell’individuo e modula
il suo livello di attività (Kay e coll., 2001; Kay e Marino, 2000). Nel
modello di “discussione cognitiva” si mette quindi in atto un
sinergismo tra la percezione cosciente dello sforzo ed il sistema
teleoanticipatorio subcosciente nella regolazione dell’intensità
dell’attività svolta (Kirkendall, 1990; Davis JM, Bailey, 1997). Un
esempio esplicativo di questo
terzo modello può essere quello di un’attività sportiva svolta in
presenza di spettatori, in questo caso l’attività stessa può
risultare spesso meno gravosa e la percezione dello sforzo minore,
proprio perché la motivazione generata dalla fonte esterna, in questo
caso gli spettatori, può ridurre gli input afferenti periferici provenienti dalla muscolatura (St Clair
Gibson e coll., 2001b). In questo caso si correre il rischio di
un’alterazione della strategia comportamentale nei confronti della
fatica, basata sull’influenza degli stimoli esterni. Infatti il
livello del meccanismo di retrocontrollo della fatica stessa, dato
dall’interazione tra il modello di discussione-cognitiva e quello
centrale-teleoanticipatorio, potrebbe elevarsi eccessivamente, e con
esso l’intensità dell’esercitazione. Il modello di
“discussione-cognitiva” quindi potrebbe essere considerato come
l’ultimo stadio d’integrazione decisionale nei confronti della
fatica, in quanto la durata e l’intensità dell’esercizio, ossia gli
aspetti decisionali nei confronti dello stesso, vengono assunti, sia in
base agli input metabolici muscolari provenienti dal modello periferico,
che secondo l’attivita centrale-teleoanticipatoria generata dal
livello corrente di attività; questi due aspetti vengono quindi
integrati nel modello di discussione-cognitiva, nel quale la percezione
della fatica proveniente dal livello di attività in corso, viene
comparata a precedenti esperienze di fatica (Kay e coll., 2001). La
fatica quindi in questo caso diviene un regolatore attivo e non più una
conseguenza passiva del processo di controllo (Kay e coll., 2001; Sargeant,
1994).
LE
MANIFESTAZIONI MIOELETTRICHE DI FATICA MUSCOLARE
Negli
ultimi anni, accanto agli studi di tipo prettamente metabolico, si sono
fortemente sviluppati dei metodi d’indagine di tipo non invasivo,
rivolti allo studio dei fenomeni bioelettrici indotti dalla fatica. I
presupposti teorici sui quali tali studi si basano sono costituiti dal
fatto che esistono particolari condizioni di lavoro durante le quali la
produzione di forza richiesta è talmente bassa da poter permettere la
prosecuzione del lavoro spesso per molte ore, questi particolari tipi di
lavoro muscolare e vengono denominati
low level static exertions. Durante questo tipo di regime di
contrazione si ipotizza che le protagoniste principali
della produzione di forza siano le unità motorie (UM) composte
da fibre di tipo ST, questa teoria
in fisiologia è nota come l’ipotesi di Cenerentola (Hägg.,
1991). Questa ipotesi è del resto del tutto conforme alla legge di
reclutamento di Henneman , secondo la quale le prime UM ad essere
reclutate e de-reclutate, in una contrazione che richieda bassi livelli
di forza, sarebbero quelle composte da fibre di tipo I. Un altro
parametro importante di cui tenere conto durante le indagini di tipo
elettromigrafico sullo studio della fatica muscolare, è costituito
dalla pressione intramuscolare (PI). Le caratteristiche del muscolo
infatti subiscono dei cambiamenti al variare della condizione ischemica
che è a sua volta correlato all’aumento della PI che si verifica
durante la contrazione muscolare stessa, soprattutto a causa della
diminuzione del flusso sanguigno distrettuale e del conseguente aumento
di metaboliti (Merletti e coll., 1984). Durante una contrazione
muscolare di tipo massimale si possono infatti raggiungere valori di PI
pari a 400-500 mmHg, mentre in contrazioni di entità molto più
modesta, compresa tra il 5 ed il 10% della massima contrazione
volontaria, il valore di PI può essere all’incirca pari a 30 mmHg. In
queste condizioni la durata della contrazione può essere mantenuta per
lungo tempo, correndo tuttavia il rischio di incorrere in una necrosi
muscolare (Sjøgard e Jensen., 1999). Tutta questa serie di fenomeni,
comporta una perturbazione del ciclo eccitazione/contrazione ed una
conseguente alterazione del segnale elettrico di superficie, nel quale
è possibile notare delle alterazioni a carico dell’ampiezza, della
forma e della velocità di propagazione del potenziale di azione. Tutta
questa serie di fenomeni è nota con il temine di “manifestazioni
mioelettriche di fatica muscolare localizzata”. Questo genere di
sperimentazioni vengono effettuate durante una contrazione di tipo
isometrico, che anche se non può essere ovviamente definita come un
pattern di attivazione perfettamente sovrapponibile al comportamento
muscolare che avviene durante una condizione naturale, offre comunque un
modello sperimentale di osservazione molto standardizzabile e
senz’altro meno influenzabile da fattori esterni non correlati al
fenomeno di fatica indagato, come ad esempio l’artefatto costituito
dal movimento del muscolo rispetto agli elettrodi di superficie che si
verifica durante un movimento dinamico (Rainoldi. e coll., 2000).
Le
variabili ed i parametri del segnale mioelettrico
Allo
scopo di caratterizzare e
rendere disponibile allo studio il segnale mioelettrico, registrato
attraverso l’elettromiografia di superficie, ossia tramite
l’applicazione di elettrodi sopra la superficie cutanea, vengono
utilizzate delle grandezze fisiche che sono classificabili “nel
dominio del tempo”, dal momento che per la propria determinazione
richiedono il solo tracciato temporale del segnale, e “nel dominio
della frequenza”, per il fatto che il loro calcolo necessiti
l’analisi spettrale del segnale e che quindi forniscano informazioni
sullo spettro del segnale stesso.
Le
variabili identificabili nel dominio del tempo, forniscono quindi
informazioni sull’ampiezza del segnale, mentre quelle appartenenti al
dominio della frequenza permettono lo studio della scomposizione in
armoniche del segnale, ossia ci danno informazioni riguardanti il
contributo in termini di ampiezza e di potenza fornito da ogni armonica
ottenuta attraverso l’analisi di Fourier del segnale stesso.
Le
variabili identificabili nel dominio del tempo, altrimenti chiamate
“variabili d’ampiezza” normalmente utilizzate nell’ambito dello
studio del segnale mioelettrico ricavato dall’elettromiografia di
superficie sono:
-
Il Valore Rettificato Medio (Average Rectified Value, ARV): che
rappresenta l’area sottesa dal segnale elettromiografico
nell’intervallo di tempo T divisa per T.
-
Il Valore Efficace (Root Mean Square), che è una grandezza
correlata alla potenza del segnale.
Le
variabili nel dominio della frequenza maggiormente utilizzate sono:
-La
frequenza media dello spettro di potenza (MNF)
, che rappresenta il valore baricentrale di frequenza dello spettro di
potenza.
-La
frequenza mediana dello spettro di potenza
(MDF), che rappresenta il valore di frequenza che divide in due
parti di eguale area lo spettro di potenza, per cui il 50% del segnale
sarà costituito da armoniche inferiori a MDF ed il restante 50% del
segnale sarà costituito da armoniche superiori a MDF.
Una
ulteriore variabile che riveste una grande importanza nello studio del
segnale elettromiografico, è la velocità di conduzione delle fibre
muscolari (CV). La CV è ricavata grazie all’utilizzo di due elettrodi
posti sulla superficie cutanea che permettono di calcolare il rapporto e/t,
intendendo con e la distanza tra i due sistemi di elettrodi e con
t
il ritardo tra il segnale registrato dal secondo elettrodo e quello
registrato dal primo. Alcuni studi hanno dimostrato come
la stima della CV sia correlata con la percentuale di fibre di
tipo II ottenibile attraverso la biopsia muscolare effettuata nel vasto
mediale di 7 marciatori e 12 sprinters (Merletti., 200). Appare quindi
chiaro l’interesse che riveste questo parametro nell’ambito di una
possibile tecnica non invasiva di determinazione della tipologia delle
fibre.
Il
cambiamento del segnale mioelettrico in condizioni di affaticamento
muscolare.
Nel corso
di una contrazione muscolare isometrica sub-massimale protratta nel
tempo, il segnale mioelettrico, a causa dell’insorgenza del fenomeno
della fatica, presenta una diminuzione della CV ed un progressivo
depauperamento dei contributi di alta frequenza. Lo spettro del segnale
si sposta verso sinistra e le armoniche significative presentano valori
progressivamente inferiori (Merletti, 2000). La diminuzione del valore
di CV comporta inoltre una concomitante diminuzione dei valori di MDF e
MNF ed un aumento dei valori di ARV ed RMS. Quindi lo spostamento verso
sinistra dello spettro del segnale e la compressione delle sue variabili riflettono il
decremento del valore di CV (Lindsrtom e Magnusson, 1977). Come
dimostrato da alcuni lavori sperimentali (Merletti e Roy, 1996), lo
studio dei cambiamenti dei parametri mioelettrici di fatica correlati
alla capacità di mantenere una contrazione isometrica sub-massimale
protratta nel tempo, possono quindi fornire importanti informazioni
sulla tipologia delle fibre muscolari considerate.

Figura 2 Spettro di potenza del segnale
elettromiografico registrato in tre distinti momenti (A, B, e C) di una
contrazione muscolare protratta nel tempo. I vari spettri sono
normalizzati rispetto al massimo valore di picco. E’ interessante
notare la diversa scala di ampiezzi dei tre spettri. (Da Merletti, 2000,
modificato)
CONCLUSIONI
Per
ciò che riguarda la fatica periferica, alla luce di questi dati, non
possiamo altro che sottolineare ancora una volta e fortemente
l’eziologia multifattoriale legata al fenomeno della sua insorgenza.
Multifattorialità quindi che esclude a priori l’esistenza di un unico
modello ma che al contrario, sottolinea l’esistenza di numerosi
fattori che si situano in altrettanto diverse e numerose tappe della
catene fisiologica esecutiva della
contrazione muscolare. Per questa ragione anche se indubbiamente la
diminuzione della concentrazione di alcuni “composti chiave” della
bioenergetica muscolare, come in particolare
la PCr ed il glicogeno, rivestono senza alcun dubbio un ruolo
chiave nel fenomeno, il ruolo della perturbazione dell’omeostasi
cellulare nella sua totalità, appare tanto determinante quanto
estremamente complesso. Potremmo comunque definire il fenomeno della
fatica periferica come un fenomeno “ a cascata” di tipo
essenzialmente protettivo, che la cellula mette in atto per preservare
la sua integrità, rispondendo all’imperativo di base di ogni
organismo vivente che altro non è che “l’autoprogrammazione per la
sopravvivenza”. Interrompere il lavoro per non autodistruggersi,
questa sembrerebbe essere
quindi la motivazione
ultima del fenomeno. Nuovi campi d’indagine, apertisi negli ultimi
anni, sembrerebbero essere particolarmente promettenti, come quelli
rivolti allo studio del ruolo dei radicali liberi, del monossido di
azoto, dell’AMP, oppure del magnesio, tuttavia una chiara ed
inequivocabile gerarchizzazione dei fenomeni che costituiscono questo
complicato, quanto perfetto meccanismo, che impedisce
l’autodistruzione cellulare, sembrerebbe ancora lontana.
Per
quello che riguarda la fatica centrale invece, se ad una parte tutta le
sperimentazioni di tipo elettrofisiologico
svolte in quest’ambito tendono unanimemente a
disegnare un suo schema di tipo lineare, che parte dalla
corteccia motrice per arrivare alla cellula muscolare, dall’altra
l’approccio di tipo neurofisiologico lascia trasparire un quadro di
notevole complessità, caratterizzato dall’integrazione di numerosi
neuromediatori, la cui funzione, se studiata isolatamente, non permette
di spiegare esaustivamente e razionalmente il fenomeno.
In ultimo
il modello tridimensionale della fatica ci permette di capire quanto i
fattori centrali siano fortemente integrati con quelli centrali di
ordine cognitivo e decisionale, sottolineando una volta di più, la
grande complessità del problema.
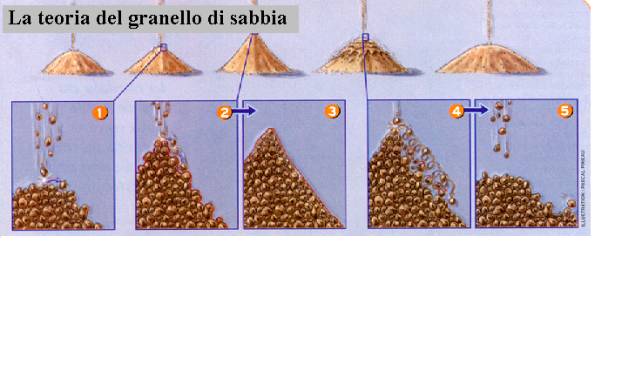
Cerchiamo quindi di capire
come avviene quello che è uno dei
disastri naturali maggiormente temuti in natura. Questo fenomeno viene
ben descritto dalla cosiddetta “teoria del granello di sabbia” che
ben illustra come il sistema raggiunga un punto di “non ritorno”
che lo porta al suo collassamento, un po’ come avviene nel nostro
organismo quando , a poco a poco, si fa strada il fenomeno della
fatica. Immaginiamo dunque un banalissimo mucchietto di sabbia, come
quello che fanno abitualmente i bambini sulla spiaggia, che cosa
succede se aggiungiamo via via dall’alto dei granelli di sabbia? In
un primo momento il nostro mucchietto diventa di dimensioni sempre
maggiori e questo sembrerebbe tutto quello che in definitiva possa
avvenire. Ma osserviamolo più attentamente da vicino: in effetti il
pendio che si viene a formare non è del tutto liscio, se avessimo a
nostra disposizione una forte lente d’ingrandimento, potremmo
facilmente notare come lungo il suo decorso si formino numerose
irregolarità costituite da tante piccolissime fossette ed altrettanto
microscopici avvallamenti, che si vengono a poco a poco a colmare con
l’aggiunta dei granelli di sabbia che cadono dall’alto.
A furia di aggiungere sabbia, la pendenza del nostro
mucchietto, ormai divenuto di una certa dimensione, è praticamente
completamente liscia, dal momento che tutte le irregolarità si sono
colmate (riquadro 3). A questo punto abbiamo raggiunto il “punto
critico”, se ora aggiungiamo ancora anche un solo granello di
sabbia, quest’ultimo non troverebbe nessuna fossa od avvallamento
dove potersi fermare e scivolerebbe inesorabilmente a valle (riquadro
4) , trascinando con se un numero più o meno importante di altri
granelli: ecco la valanga. Dopo l’evento del fenomeno, ossia dopo la
discesa a valle della valanga, il cumulo di sabbia, ma a questo punto
potremmo dire anche di neve, ritorna nuovamente irregolare come
all’inizio ed un nuovo ciclo può compiersi.
BIBLIOGRAFIA
Allen
DG., Lamergren J., Westerblad H. Muscle cell function during
prolonged activity: cellular mechanism of fatigue. Exp.
Physiol. 80: 497-527, 1995.
Atlan G.,
Beliveau I., Bouisson P. La fatigue musculaire. Muscle fatigue.
Masson Ed. Paris, 1991.
Bangsbo J., Gollink PD., Graham TE., Jael C., Kiens B., Mizumo M et al. Anaerobic
energy production and O2 deficit-debt relationship during exhaustive
exercise in humans.
J. Physiol. 422: 539-559, 1990.
Bassett DR, Howley ET. Maximal oxygen uptake: "classical"
versus "contemporary" viewpoints. Med Sci Sports Exerc.
29:591-603, 1997.
Bayley SP., Davis JM., Ahlborn EN. Brain
serotoninergic activity affected endurance performance in the rat. Int
J. Sport Med.6: 330-333, 1993.
Bigland-Ritchie B., Jones DA.,
Woods J. Excitation frequency and muscle fatigue: electrical reponse
during human voluntary and stimulte contraction. Exp Neurol 64:
414-427, 1979.
Blomstrand
E., Hassmen P, Newsholme E. Effect of branched chain aminoacids
supplementation on mental performance.Acta Physiol Scand. 136:
473-481, 1991.
Chauloff F. Physical Exercise and brain monoamines:
a review.Acta Physiol Scand. 137: 1-13, 1989.
Chin
ER., Allen DG. The
contribution of pH-dependent mechanism to fatigue at different
intensities in mammalian single muscle fibres. J. Physiol. 3:
831-840, 1998.
Danieli
D., Germinaro E., Esposito A., Biral D., Betto R. Effects of fatigue
on sarcoplasmatic reticulum and myofibrillar properties of rat slow and
fast twitch muscle fibers. J Appl Physiol. 89: 891-898, 2000.
Davis JM, Bailey SP. Possible mechanisms of
central nervous system fatigue during exercise. Med Sci Sports Exerc.
29:45-57, 1997.
Dishman RK. Brain monoamides exercise and
brhaviour stress: animal models. Med Sci Sports Exerc. 29: 63-74,
1997.
Enoka RM., Stuard DG. Biology of muscle fatigue.
J Appl Physiol. 72: 1631-1648, 1992.
Fitts
H. Metzger JM. Mechanism
of muscular fatigue. In: Poortmans JR, ed Principles of exercise
biochemistry, 2nd, rev. ed. Med.
Sport. Sci. Basel. Karger 38: 248-268, 1993.
Fitts
H. Muscle fatigue : the cellular aspects. Am.
J. Sports. Med.
6: S9-s13, 1996.
Fitts RH. Cellular mechanisms of muscle fatigue.
Physiol
Rev.74:49-94, 1994.
Gandevia SJ. Allen GM.,
Butter GE., Taylor GL. Supraspinal factor in human muscle fatigue. Evidence for subspinal output
from thr motor cortex. J Physiol. 490: 529-536, 1996.
Green
HJ. Mechanism of muscle
fatigue in intense exercise. J. Sport. Sci. 15: 247-256, 1997.
Green
HJ. Cation pump in skeletal muscle: potential role in muscle fatigue.
Acta. Physiol. Scand. 3: 201-214, 1998.
Greenhaff
PL. Creatine and its application as an ergogenic aid. Int J Sport
Nutr. 5 suppl. S100-S110, 1995.
Guézennec CY. La
fatigue centrale: électrophysiologique ou neurochimique. Science
& Sport. 15 : 228-233, 2000.
Hägg.
G. Static work loads and occupational myalgia – a new explanation
model. In: PA. Anderson, DJ. Hobart, JV. Danoff (Eds). Electromyographical Kinesiology (Elsevier
Science, Amsterdam), 141-143, 1991.
Hirvonen
J., Nummela A., Rusko H., Rehunen S. Haerkoenen M. Fatigue and
changes of ATP, creatine phosphate, and lactate during the 400 m sprint.
Canadian Journal of Sport Sciences. 17(2): 141-144, 1992.
Hirvonen
J., Rehunen S., Rusko H.,
Haerkoenen M. Breakdown of hight energy phosphate compounds and
lactate accumulation during short supramaximal exercise. Eur J Appl
Physiol and Occ Physiol. 56(3): 253-259, 1987.
Hogan
MG., Bruce Gladden L., Kurdak SS., Poole DC. Increased lactate in
working dog muscle reduces tension development independent of pH. Med
Sci Sport Exerc. 3: 371-377, 1995.
Jacobsen
BJ., Coast R., Donnelly JE. The
effect of exercise intensity on slow component of VO2 in
person of different fitness level. J Sport Med Phys Fitness. 38: 1245-131, 1998.
Kay D, Cannon J,
Marino FE, St Clair Gibson A, Lambert MI, Noakes TD. Evidence for
neuromuscular fatigue during cycling in warm humid conditions. Eur
J Appl Physiol. 84:115-121, 2001.
Kay D, Marino FE. Fluid
ingestion and exercise hyperthermia: implications for performance,
thermoregulation, metabolism, and the development of fatigue.
J
Sport Sci. 18:71-82, 2000
Kirkendall DT. Mechanisms of peripheral
fatigue. Med Sci Sports Exerc. 22:444-449, 1990.
Korge
P., Campbell KB. The importance of ATPase microenvironment in muscle
fatigue: a hypothesis. Int. J. Sports Med. 3: 172-179, 1995.
Koutedakis Y., Frischknecht R., Vrova G., Sharp G.,
Budgett R. Maximal voluntary quadriceps strenght patterns in olympic
overtrained athletes. Med
Sci Sports Exerc. 27: 566-572, 1995.
Linde PK., Verburg E., Vellestad N., Sejersted OM.
Skeletal muscle fatigue in normal subjects and heart failure patients. Is
there a common mechanism ? Acta. Physiol. Scand. 3: 215-228, 1998.
Linderman
JK., Gosselink KL. The
effects of sodium bicarbonate ingestion on exercise performance.
Sports. Med. 2: 75-80, 1994.
Lindsrtom
L., Magnusson R. Interpretation
of myoelectric power spectra: a model and its application. Proc.
IEEE, 65: 653-662, 1977.
Maione D., Senaldi R.,
Gnudi G., Maietta P., Tentoni C., Drago E. La nostra esperienza
nell’utilizzo della cinetica del VO2 per il monitoraggio
dell’allenamento fisico. Medicina dello Sport. 54:
51-62, 2001.
Mannion
AF., Jakeman PM., William PL. Skeletal
muscle buffer value, fibre type distribution and high intensity exercise
performance in man.
Exp. Physiol. 80: 89-101, 1995.
Maton B. Central nervous changes induced by local
work. In Atlan G., Belivau L., Boiussou P. Ed. Muscle
fatigue. Paris. Masson. p 207-222, 1991.
Mc
Lester JR. Muscle contraction and fatigue. The role of adenosin5’-diphosphate
and inorganic phosphate. Sport. Med. 5: 287-305, 1997.
Meeusen R., De Meirlir K. Exercise and brain neurotransmission. Sports
Med. 20: 160-188, 1995.
Merletti
R., Sabbahi MA., De Luca CJ., Median frequency of the myoelectric
sygnal – Effects of muscle ischemia and cooling. Eur
J Appl Physiol 52: 258-265, 1984.
Merletti R. Elementi
di elettromiografia di superficie. C.L.U.T (Eds), Torino, pg 55-62, 2000.
Merletti
R., Roy S. Myoelectric and mechanical manifestation of muscle fatigue
in Voluntary contractions. JOSPT, vol 24, 6: 342-353, 1996.
Mujika
I., Padilla S Creatine supplementation as an ergogenic acid for sport
performance in highly trained athletes: a critical review. Int J
Sport Med. 18: 491-496, 1997.
Newsholmes
EA., Acworth IN., Bloomstrand E. Aminoacids brain neurotransmission
and a functional link between muscle and brain that is important during
sustained exercise. In: G. Benzi Ed. Advances
in myochemistry. London: John Libey Eurotext Ltd.p 127-138, 1987.
Nielsen OB., Harrison AP. The
regulation of Na+.
K+ pump in
contracting skeletal muscle.
Acta Physiol. Scand. 3: 191-200, 1998.
Nielsen
OB., Overgaard K. Ion
gradients and contractility in skeletal muscle: the role of active Na+ .
K+ transport. Acta. Physiol: Scand. 156: 247-256, 1996.
Paterson
DL. Role of potassium in the regulation systemic physiological
function during exercise. Acta Physiol Scand.
156: 287-294, 1996.
Perronet
F. Significations et limites de la lactacidémie dans le contrôle de
l’entraînement, Actes du Troisième Colloque International de la
Guadeloupe, 209-221, 1994.
Pybus
J, Tregear RT. The relationship of adenosine triphosphatase activity
to tension and power
output of insect flight muscle. J Physiol. 247(1):71-89, 1975.
Rainoldi
A., Nazzaro M., Merletti R., Farina D., Caruso I., Gaudenti S. Geometrical
factors in dynamic surface EMG of the vastus medialis and lateralis. J
of Electrom and Kinesiol. Vol 10 n° 5: 327-336, 200.
Rios E., Ma J., Gonzales A. The mechanical hypotesis of
excitation-contraction (EC) coupling in skeletal muscle. J Muscle Res
Cell Motil. 12: 127-135, 1991.
Rios
E., Pizzarro G. Voltage sensor and calcium channels of
excitation-contraction coupling. N Physiol Sci. 33: 223-227, 1988.
Rohmer TW. Die bezichung zwischen kraft und ausdeuer bei statisher
muskelabeirt, schriftenreihe arbeitsmedizin, sozialmedizin.
Arbeitshygiene band 22, AW. Stuttgard: Gebtber-Verlag; 118, 1968.
Rouillon JD., Candau R. La fatigue périphérique : sites
subcellulaires et mécanismes biologiques. Science
& Sport 15 : 234-241, 2000.
Sahlin
K. Acid-base balance during high intensity exercise. In: Hatties
M., Williams C., Stanish WD., Micheli LG., Eds. Oxford textbook of sport
medicine. Oxford University Press. 1994, p 46-52.
Sahlin
K. Metabolic aspects of fatigue in human skeletal muscle. In:
Muscle fatigue mechanism in exercise training: Marconnet P Ed.
Proceedings of the 4th International Symposium on
Exercise and Sport Biology. Basel
S. Karger Publisher. 54-68, 1992.
Sahlin K., Ren JM. Relationship of contraction capacity to metabolic changes during recovery
from a fatiguing contraction.
J, Appl. Physiol. 67(2): 684-654, 1989.
Sahlin
K., Tonkonogi M., Söderlund K. Energy supply and muscle fatigue in
humans. Acta Physiol Scand. 162: 261-266, 1998.
Sargeant
AJ. Human power output and muscle fatigue. Int J Sports Med.
15:116-121, 1994.
Seguin L., Liscia P., Guézennec CY., Fillion G. Effect
of moderate and intensive training on functional activity of central 5HT
1b receptors in the rat substantia nigra. Acta Physiol Scand. 162:
63-68, 1998.
Sejersted
OM, Vallestad NK, Hallein J., Bahr R, Muscle performance-fatigue,
recovery and trainability. Acta Physiol Scand. 3: 181-182, 1998.
Shepard
RJ. Physiology &.biochemistry
of exercise. Praeger Ed. New York, 1986.
Sjogaard
G. Potassium and fatigue: the pros and cons. Acta Physiol Scand.
156: 257-264, 1996.
Sjøgard
G., Jensen BR., Low
level static exertions. In: The occupational ergonomic handbook. CRC
Press, 1999.
Spriet LL, Soderland K, Bergstrom M, Hultman E. Anaerobic energy
release in skeletal muscle metabolism during electrical stimulation in
men. J Appl Physiol. 62:611-615, 1987.
St Clair Gibson A, Schabort EJ, Noakes TD. Reduced
neuromuscular activity and force generation during prolonged cycling.
Am J Physiol Regul Integr Comp Physiol. 281:R187-R196, 2001.
St
Clair Gibson A., Lambert EV., Hampson D., Noakes TD. Exercise and
fatigue-control mechanism. International Sport Med J. 2(3): 54,56, 2001
Stephenson
DG., Lamb GD., Stephenson GMM. Events of the
excitation-contraction-relaxation (E-C-R) cycle in fast and slow-twitch
mammalian muscle fibres relevant to muscle fatigue. Acta
Physiol Scand. 3: 229-246, 1998.
Ulmer H-V. Concept of an extracellular regulation of muscular
metabolic rate during heavy exercise in humans by psychophysiological
feedback.Experentia. 52:416-420,1996.
Wagenmakers
AJ. Role of amino acids and ammonia in mechanisms of fatigue. In:
Marconnet P,
Komi
PV, Saltin B, Sejersted OM, eds. Muscle Fatigue Mechanisms in Exercise
and Training. Basel,
Switzerland: Karger; 34:69-86. Medicine and Sport Science; Vol. 34, 1992.
Westerblad H., Lee JA., Lamb AG., Bolsover SL., Allen DG. Spatial
gradients of intracellular calcium in skeletal muscle during fatigue.
Pfulgers Arch. 6: 734-740, 1990.
Westerblad H., Lee JA., Lannergren J., Allen DG. Cellular
mechanism of fatigue in skeletal muscle.
Am. J Physiol. 261:C195-C209,
1991.
Whipp
BJ., Wassermann K. Oxygen uptake kinetics for various intensities of
costant-load work. J Appl Physiol33: 351-356, 1970.
Williams
JH., Klug GA. Calcium exchange hipothesis of skeletal muscle fatigue:
a brief review. Fatigue and Calcium Exchange Muscle Nerve. 18:
421-434, 1995.
Wooledge
RC. Possible effects of fatigue on muscle efficiency. Acta
Physiol Scand. 3: 267-274, 1998.
ATPasi : Sigla di
Adenosintrifosfatasi, enzima che catalizza l'idrolisi dell'adenosintrifosfato
(ATP) in adenosindifosfato (ADP) e fosfato (Pi). L'idrolisi dell'ATP
si accompagna sempre ad un altro processo biochimico o fisiologico.
Le ATPasi della membrana citoplasmatica e del reticolo
sarcoplasmatico, per esempio, funzionano da pompe ioniche; la ATPasi
miosinica permette la conversione dell'energia chimica in energia
muscolare, ecc. Alcune ATPasi catalizzano, in vivo, la reazione
inversa; è questo il caso della ATPasi mitocondriale, che
sintetizza ATP sfruttando il gradiente protonico esistente ai due
lati della membrana mitocondriale interna.
Corpo striato: Nella
neuroanatomia dei Mammiferi descrive lo striato telencefalico.. Il
corpo striato rappresenta prevalentemente una stazione intercalata
sulle vie corticofugali extrapiramidali ma sono descritte anche
connessioni a doppio senso con il talamo dorsale.
|